Proyecto Titi has been actively involved in studying wild cotton-top tamarins for many years at our field sites in Colosó, Santa Catalina, and San Juan Nepomuceno. We present a short overview of some of our publications on the behavior, reproductive patterns, and parental care strategies of this fascinating primate. As habitat for cotton-tops continues to be threatened in Colombia, it is essential that we document how this will effect this critically endangered primate for the future. We will continue to post information on our publications as they become available.
Cotton-top tamarins (Saguinus oedipus) are a critically endangered species found only in Colombia. Their survival is threatened by extensive habitat destruction and the illegal pet trade. Because many people in Colombia have a low level of awareness of cotton-top tamarins and even less understanding of the impacts that the illegal pet trade has on the species and its long-term survival, Proyecto Tití has developed a series of programs for children in rural elementary schools that introduce children to cotton-top tamarins. This study examines Proyecto Titi's efforts to reduce the desire to have a cotton-top tamarin as a pet with outcomes focused specifically on (1) creating an emotional connection between people and cotton-top tamarins, (2) identifying the feelings and understanding the challenges that cotton-top tamarin face when they are kept as pets, and (3) empowering students to make positive choices to help cotton-top tamarins. Both TITI KIDS and TITIRITIANDO programs helped students to (1) increase their knowledge of cotton-top tamarins, (2) understand the damage that the pet trade has on individual animals and the long-term survival of cotton-top tamarins and (3) provide practical actions that they can engage in to protect cotton-top tamarins for the future. Our hope is that students from our programs become advocates for conservation in their communities and assist in the long-term conservation activities to protect cotton-top tamarins in Colombia.
Maynard, L., Savage, A., Vega, J., DeWan, A., Diaz, L., Gezon, Z., Guillen, R. 2021. Can creating sustainable livelihoods with communities impact cotton-top tamarin (Saguinus oedipus) conservation in Colombia? Conservation Science and Practice, 3(8) e476.
Conservation and development organizations around the world are increasingly promoting livelihood programs for rural communities as a means of creating sustainable alternatives to activities destructive to remaining wildlife and habitats. In impoverished communities in Colombia adjacent forest patches with critically endangered cotton-top tamarins (Saguinus oedipus), this study evaluates alternative livelihood programs' impact on participating community members' income, wellbeing, sustainable resource use, and attitudes about cotton-top tamarin conservation. We surveyed program participants, their neighbors, and members of reference communities nearby for comparison (n = 253). Participants in the alternative livelihood programs had significantly more access to resources and benefits compared to both the non-participating neighbors and people from the reference communities. However, results revealed challenges of scaling up the programs to include more community members. Participants' attitudes toward sustainable resource use and cotton-top tamarin conservation were not stronger than others who do not receive benefits from the alternative livelihoods programs, and at times were contrary to the conservation goals, revealing a gap in program training for newer participants. Conservation programs using alternative livelihood strategies can apply the lessons learned in our 16 years of implementation and this post hoc evaluation to strengthen the connection between alternative livelihoods and conservation goals.
Feilen, K.L., Guillen, R.R., Vega, J., Savage, A. 2018. Developing successful conservation education programs as a means to engage local communities in protecting cotton-top tamarins (Saguinus oedipus) in Colombia. Journal for Nature Conservation, 41, 44-50.
Engaging children in developing countries to care for wildlife, and to make appropriate choices to protect wildlife, are challenging tasks. Proyecto Tití works to conserve the cotton-top tamarin (Saguinus oedipus), a critically endangered endemic monkey from Colombia, using a multi-disciplinary approach that focuses on scientific research, community empowerment, habitat protection, and conservation education to protect this species. Proyecto Tití has developed a multi-tiered methodology for education in rural communities that increases knowledge and understanding of the cotton-top tamarin and the threats to its survival, generates attitudes and behavior that support conservation, and provides opportunities to develop the future generation of conservation leaders in Colombia. We evaluated TITÍ KIDS and CARTITILLA, two of Proyecto Tití’s educational programs, measuring the knowledge gained by students before and immediately after the program. For CARTITILLA, we also measured the retention of knowledge after five years, and if the program developed pro-conservation attitudes and behaviors. We analyzed the evaluations of over 6000 students who participated in programs from 2011 to 2016, using a paired t-test. Students significantly increased their knowledge of cotton-top tamarins (p ≪ 0.01) and their ability to identify and classify domesticated and wild animals in their regions (p ≪ 0.01). Five years after participating in the CARTITILLA program, students had retained their knowledge about cotton-top tamarins and demonstrated attitudes and behavior that support cotton-top tamarin conservation. The educational programs of Proyecto Tití provide evidence that engaging youth in meaningful conservation activities can assist in the protection of cotton-top tamarins.
Savage, A. & Guillen, R. 2012. Conserving cotton-top tamarins Saguinus oedipus through effective captive management, pubic engagement, and in situ conservation efforts. International Zoo Yearbook 46: 56-70.
The Cotton-top tamarin Saguinus oedipus is a Critically Endangered primate found only in Colombia. While a popular exhibit animal in zoos and an important research subject in the study of colon cancer and reproductive biology and behaviour, efforts to create a self-sustaining captive-breeding population for this species have been very successful. With the wealth of information on biology, captive management and status in the wild, efforts are now focused on ensuring that Cotton-top tamarins continue to remain in the wild. With large-scale development threatening some of the last remaining forested areas for the species and its popularity in the illegal pet trade, compounded by the constant extraction of forest resources by impoverished communities, strategies are needed to protect this species effectively for the future. Here, an overview is presented of the successful, multidisciplinary conservation programme developed by Proyecto Tití to involve local communities, national and international agencies, and the conservation community in the long-term protection of Colombia’s most threatened species.
DEMOGRAPHY AND GROUP COMPOSITION
Savage A, Giraldo L.H, Blumer E.S, Soto L.H, Snowdon C.T. 1996. Demography, group composition, and dispersal in wild cotton-top tamarin (Saguinus oedipus) groups. American Journal of Primatology 37:23-32.
Tamarins live in social groups of 2-10 individuals. Although, there have been a few cases of reported polygamy, most groups contain only one reproductively active male and female (Neyman 1977). Females, on average, give birth to twins annually. Social groups appear relatively stable, with an average rate of emigration of 0.71 ± 1.18 individuals/group/year. Males and females disperse to neighboring groups equally, however, adults move more frequently than juveniles and infants. Immigrant males are more likely to enter a group following the death of a resident male. Reproductive success of these immigrant males is difficult to quantify without paternity tests, however, an immigrant male may release subordinate females from reproductive suppression. A novel male avoids the constraints of incest avoidance, and may thus stimulate a reproductively inhibited female. If a female immigrates, she does not necessarily assume the breeding position. Emigration increased to 2.8 individuals/group/year during a drought , suggesting movement between groups may be a result of extreme environmental conditions leading to increased competition for limited resources.
A proposed model to illustrate the potential reproductive strategies of cotton-top tamarins:
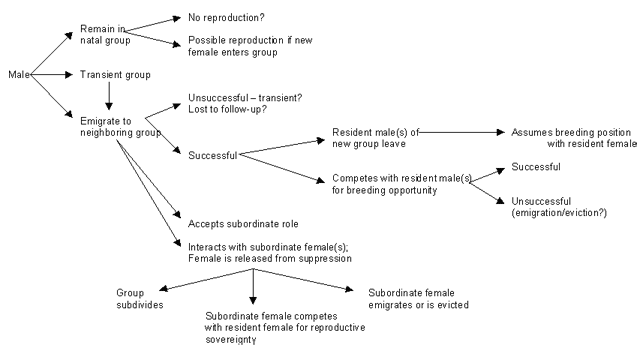
Return to Top
DIET - PLANT SPECIES CONSUMED BY COTTON-TOP TAMARINS IN COLOMBIA
Savage, A., Snowdon, C.T., Soto, L., Castro, J., Medina, F., Emeris, G., Garcias, L.C., Guillen, R. 2022. The impacts of seasonal variation and climate on food utilization in a population of critically endangered cotton-top tamarins (Saguinus oedipus) in Colombia: A 22 year longitudinal study. American Journal of Primatology, DOI: 10.1002/ajp.23415
To examine how precipitation patterns and climate change impact feeding choices made by a population of critically endangered cotton-top tamarins (Saguinus oedipus), we examined 22 years of feeding data (1999–2020) from 21 groups collected at Parque Natural Regional Bosque Seco El Ceibal Mono Tití in Santa Catalina, Colombia. We describe the diet and examine the role of seasonal rainfall and annual variation in rainfall on diet. Rainfall is highly seasonal (mean annual rainfall 1562 mm [range 940–2680 mm]) with a dry, early rainy, and late rainy season in each year. Over 80 species of plants formed part of the fruit, nectar, and exudate components of the diet. Fruits, although available year-round, were more commonly available and consumed during the late rainy seasons (August–November). Exudates were consumed more frequently in the dry season (December–March) and invertebrate consumption was stable across the year. Nectar feeding from a single species (Combretum fruticosum) peaked in November. Rainfall varied over the years, with 13 years exceeding the 99% confidence intervals for mean rainfall. Ten of these extreme years (both drought and extremely wet) occurred in the last 11 years. Fruit consumption did not vary between extreme and average years, but cotton-top tamarins consumed more invertebrates and exudates in wet years. Presently, cotton-top tamarins appear to be able to cope with these extreme variations in rainfall due to their highly varied diet. However, the forests that these primates depend upon for survival are threatened by human exploitation making it critically important to maintain a generalist feeding strategy for survival as many fruiting trees that compose a large proportion of the diet are removed. As conservation efforts continue, plant species consumed by cotton-top tamarins provide useful data when selecting species for habitat restoration programs.
Return to Top
FIELD TECHNIQUES
Ruiz Miranda, C.R., Savage, A., Forero, F. 2023. Aplicaciones del monitoreo acústico activo y pasivo para conservación de New World Primates. Neotropical Primates 29(1) 1-13.
Most Neotropical primates are very vocal, and many species use long–distance calls for intra and inter–group communication. The use of field methods based on acoustic signaling holds promise for Neotropical primate population studies when combined with remote sampling techniques. However, if assumptions are not met, these methods may be ineffective or provide misleading results. We will address the use of passive and active acoustic monitoring (playback)for the survey and monitoring of primate populations. We discuss the suitability of each method in terms of the target species and project objectives, data analysis approaches, team considerations, research design, and other factors affectingprimate detection in the field.
Savage, A., Giraldo, L.H., Blumer, E.S., Soto, L.H., Burger, W., and Snowdon, C.T. 1993. Field techniques for monitoring cotton-top tamarins (Saguinus oedipus oedipus) in Colombia. American Journal of Primatology 31: 189-196.
We present reliable field techniques for capturing, anesthetizing, and identifying individual cotton-top tamarins (Saguinus oedipus oedipus). A new technique is presented for radio-tracking small bodied primates. A backpack-style harness was designed to carry a transmitter. This system appears effective in minimizing potential injury and does not appear to interfere with the normal behavior of the animal.
Return to Top
GENETICS
Rasmussen, L. Fontsere, C., Soto-Calderon, I.D., Guillen, R., Savage, A., Hansen, A.J., Hvilsom, C., Gilbert, T.P. 2023. Assessing the genetic composition of cotton-top tamarins (Saguinus oedipus) before sweeping anthropogenic impact. Molecular Genetics
During the last century, the critically endangered cotton‐top tamarin ( Saguinus oedipus ) has been threatened by multiple anthropogenic factors that drastically affected their habitat and population size. As the genetic impact of these pressures is largely unknown, this study aimed to establish a genetic baseline with the use of temporal sampling to determine the genetic makeup before detrimental anthropogenic impact. Genomes were resequenced from a combination of historical museum samples and modern wild samples at low‐medium coverage, to unravel how the cotton‐top tamarin population structure and genomic diversity may have changed during this period. Our data suggest two populations can be differentiated, probably separated historically by the mountain ranges of the Paramillo Massif in Colombia. Although this population structure persists in the current populations, modern samples exhibit genomic signals consistent with recent inbreeding, such as long runs of homozygosity and a reduction in genome‐wide heterozygosity especially in the greater northeast population. This loss is likely the consequence of the population reduction following the mass exportation of cotton‐top tamarins for biomedical research in the 1960s, coupled with the habitat loss this species continues to experience. However, current populations have not experienced an increase in genetic load. We propose that the historical genetic baseline established in this study can be used to provide insight into alteration in the modern population influenced by a drastic reduction in population size as well as providing background information to be used for future conservation decision‐making for the species.
Return to Top
HEALTH
Power, M.L., Oftedal, O.T., Savage, A., Blumer, E.S, Soto, L.H., Chen, T.C., Holick, M.F. Assessing Vitamin D status of Callitrichids: Baseline data from wild cotton-top tamarins (Saguinus oedipus) in Colombia. Zoo Biology, 16:39-46, 1997.
Bone disease related to vitamin D deficiency is an insidious problem in captive colonies of callitrichids. Currently a diagnosis of vitamin D deficiency may be made after the appearance of clinical pathology, usually bone deformities or fractures. The development of assays for vitamin D metabolites suggests it may be possible to detect incipient vitamin D deficiency before animals are adversely affected. However, there are few data on normative levels of these metabolites in any nonhuman primates. We collected blood samples from 18 wild cotton-top tamarins (Saguinus oedipus) from Colombia and assayed them for 25-hydroxy vitamin D(25-OH-D), the major circulating form of the vitamin, which is believed to be a good indicator of status. Serum concentrations of 25-OH-D averaged 76.4 ng/ml (range 25.5–120 ng/ml). This is high compared with human norms (10–55 ng/ml), but the range is lower than that reported in the literature for captive callitrichids. Juveniles had higher serum concentrations than did adults, and pregnant females had lower concentrations than did nonpregnant females.These data confirm that healthy callitrichids have higher circulating levels of 25-OH-D than do humans, but they suggest that the extremely high levels found in some captive animals (300–600 ng/ml) may be above normal. We propose that serum concentrations of 25-OH-D of 50–120 ng/ml can be considered normal for cotton-top tamarins and perhaps other callitrichids. If serum values much below 50 ng/ml are found during clinical evaluation, the possibility of incipient vitamin D deficiency should be considered.
Return to Top
INFANT DEVELOPMENT AND PARENTAL CARE
Ziegler, T.E., Tecot, S.R., Fernández-Duque, E., Savage, A., Snowdon, C.T., 2022. Nonhuman primate paternal care: Species and individual differences in behavior and mechanisms. In: Patterns of Parental Behavior, Ed. G. Gonzáles-Mariscal, Springer, https://doi.org/10.1007/978-3-030-97762-7_7
Direct care of offspring by the father (sire) is relatively rare in primates. Besides humans, there are a number of species where the male is essential for the survival of offspring: marmosets, tamarins, titis and owl monkeys, some lemurs, and siamangs. All these species show reduced sexual dimorphism, territoriality, and biparental care. However, timing and levels of direct care may vary among these species. Here, relying on both lab and field data, we address the variability found in father's involvement with his infants, the behavioral, neuroendocrine and sensory systems that are a cause and consequence of paternal care, and social bonds between the breeding pair. We integrate studies of laboratory animals (where detailed observations and experimentation are possible) with field studies (which illuminate the ecological and evolutionary functions of paternal care) and discuss the future directions for examining the proximate and ultimate mechanisms of paternal care in nonhuman primates.
Ziegler, T.E., Tecot, S.R., Fernández-Duque, E., Savage, A., Snowdon, C.T., In press. Nonhuman primate paternal care: Species and individual differences in behavior and mechanisms. Advances in Neurobiology. 2022;27:213-238. doi: 10.1007/978-3-030-97762-7_7.
Direct care of offspring by the father (sire) is relatively rare in primates. Besides humans, there are a number of species where the male is essential for the survival of offspring: marmosets, tamarins, titis and owl monkeys, some lemurs, and siamangs. All these species show reduced sexual dimorphism, territoriality, and biparental care. However, timing and levels of direct care may vary among these species. Here, relying on both lab and field data, we address the variability found in father’s involvement with his infants, the behavioral, neuroendocrine and sensory systems that are a cause and consequence of paternal care, and social bonds between the breeding pair. We integrate studies of laboratory animals (where detailed observations and experimentation are possible) with field studies (which illuminate the ecological and evolutionary functions of paternal care) and discuss the future directions for examining the proximate and ultimate mechanisms of paternal care in nonhuman primates.
Savage, A., Snowdon, C.T., Soto, L., Medina, F., Emeris, G., Guillen, R. 2021. Factors influencing the survival of wild cotton-top tamarin (Saguinus oedipus) infants. American Journal of Primatology, 83(7) e23262
Studies of cooperative breeding species have suggested that helpers are needed for infant survival and that helpers gain skills to successfully raise their own offspring. Studies of callitrichids in managed care and early field studies suggested that group size correlated with infant survival and that helpers needed to learn parental skills to be successful breeders. We present infant survival data from a 20-year field study of cotton-top tamarins (Saguinus oedipus) in Colombia involving 126 litters born to 41 females. There was no difference in the survival of male and female offspring to 6 months of age. However, litter size impacted survival, with triplet litters having lower survival and male triplets having lower survival than females. Loss of infants was highest in the first week of life and, of the animals remaining in the group at 6 months of age, 90% of them survived to 1 year. The mean length of time in the natal group was 2.3 years but some young migrated to other groups after 4 months of age. Group size and number of male helpers (>1 year) was not related to infant survival. However, groups with only one male had fewer surviving infants. Primiparous females had lower infant survival, but previous infant care experience by mothers as helpers had no effect on survival. A major cause of infant loss was group disruption due to deaths/evictions/loss of the reproductively active animals, or immigration of pregnant females. Thus, factors that cause a change in the stability of the reproductively active animals can disrupt group cohesion. Capture of individuals for the illegal pet trade, as well as habitat fragmentation, may increase competition between groups for access to territories and breeding opportunities. This may have long-term impacts to infant survival in this critically endangered species.
Savage, A., Soto, L., Medina, F., Emeris, G., & Soltis, J. 2009. Litter Size and Infant Survivorship in Wild Groups of Cotton-top Tamarins (Saguinus oedipus) in Colombia. American Journal of Primatology, 71, 1-5.
Developing successful conservation programs for endangered primates requires a complete understanding of the factors involved in successful reproduction and causes of infant mortality. In contrast to other nonhuman primates, callitrichids are unusual as litter size can vary between one to five offspring in captivity. Twins tend to be the norm for most callitrichids, but the incidence of triplet and quadruplet births can account for nearly 10–50% of the litters born in captive colonies. Litter size significantly influences the survivorship of infants with twins having a higher survivorship than triplets or quadruplets. In the wild, reports suggest that twin and singleton litters were the norm in most callitrichids studied to date. Incidence of triplet births is very low in the wild, but there have been triplets observed in a Golden Lion Tamarin family where none of the offspring survived and in a Common Marmoset family where evidence was found of survival.
In our study, there were fifty-eight litters from 21 females recorded at the two field sites. Twins were most common (81%), followed by singletons (16%), and we had the first report of triplet births (3%) in wild cotton-top tamarins. Mortality was not uniformly distributed across time with 33% of cases confined to the first week. Determining the sex of the offspring rarely occurred if infants died or were missing from the group, and carcasses of infants were rarely found. Thus, calculating an accurate sex ratio of infants was not possible. However, surviving offspring were comprised of 42 males and 32 females. Determining the immediate cause of mortality was also challenging. Two of the six carcasses that were recovered were too decomposed to determine the cause of death. The remaining four carcasses were from the two observed triplet births and malnutrition was deemed the cause of death.
Fourteen females produced more than one litter. For these females, the mean survival rate to 6 months for first litters was 57%710.3 SEM but the mean across subsequent litters rose to 86%77.6. However, the effects of parity were not as clear. One primiparous female (Nadia) left her family group and assumed the position of the reproductively active female in a neighboring group and she gave birth to twins that survived to 6 months. However, two primiparous females (Milena and Tamara) were identified in the Santa Catalina sample but do not represent ‘‘typical’’ primiparous females as they were in groups containing two pregnant females. Milena successfully cared for her twins until one was observed missing from the group at week 8 and Tamara’s twins were ‘‘kidnapped’’ by her mother and subsequently died. Litter size appeared to influence survival with triplet litters showing a lower infant survival to 6 months (33%) than twins (80%) or singletons (77%), although the number of triplet litters (n52) is too small for statistical analysis.
This study confirms that infant survival to 6 months of age in the wild (78% twins and 88% singletons), gathered from two distinct field sites, and was greater than has been reported in captive colonies. Similar to captive colonies mortality was highest during the first week of life; however, given the low rate of carcass retrieval we cannot determine the cause of mortality and infant mortality associated after 20 days may be related to the development of infant independence. Given that the parity of the majority of our study females could not be determined, we did find that infant mortality was higher in first litters than subsequent litters born to wild cotton-top tamarin females. We were unable to find a significant relationship between infant mortality and group size or the number of adult males in a social group. However, there was a trend toward higher mortality with a shorter inter-birth interval. The number of females that produce multiple litters in 1 year is rare, but infants born out of the normal birthing season may be at risk as food may not be as plentiful for a nursing female.
Savage A. Snowdon CT, Giraldo LH. 1996b. Parental care patterns and vigilance in wild cotton-top tamarins (Saguinus oedipus). In Adaptive Radiations of Neotropical Primates. Eds. M Norconk, A Rosenberger, P Garber. New York: Plenum Press.
Parental care in this species is shared by all group members. Early infant caretaking experience, observed in captivity, has been found to influence future reproductive success in both males and females. If an animal has never carried an infant on its back while it was in its family it will abuse its own offspring. However, if the animal has experience caring for offspring prior to reproduction, it will successfully care for its offspring. Interestingly, non-natal animals are actively involved in infant care and are often observed assuming sentinel positions in their new groups. An animal never carries infants while involved in “sentry” duty. The sentry remains vigilant while the group forages or rests and is responsible for notifying the group of any potential threats to their safety.
Observations of 12 births in the wild showed that infants are carried exclusively during the first four weeks of development, with a gradual decrease during weeks 5-9, such that by week 10, the infants are solo for nearly 50% of the observation time. All individuals in the group carry the infants, though adults were more likely to than juveniles. There was no significant difference in carrying time between males and females. Primiparous females, however, carry their infants more during their first two weeks, than multiparous females. Infant survival to one year of age increases with group size (see Table 1 below). Contribution of the mother in infant care remains independent of group size, however, the larger the group, the less time the adult males carried infants.
| Group Size (N) |
Infant Survival Rate |
| 3 |
40% |
| 4 |
66.7% |
| 5 |
100% |
| 6 |
87.5% |
Return to Top
POPULATION AND HABITAT ASSESSMENT
Savage, A., Thomas, L., Feilen, K.L., Soto, L.H., Pearson, M., Medina, F.S., Emeris, G., Guillen, R.R. 2016. An assessment of the population of cotton-top tamarins (Saguinus oedipus) and their habitat in Colombia. PLoS One 11 (12) e0168324.
Numerous animals have declining populations due to habitat loss, illegal wildlife trade, and climate change. The cotton-top tamarin (Saguinus oedipus) is a Critically Endangered primate species, endemic to northwest Colombia, threatened by deforestation and illegal trade. In order to assess the current state of this species, we analyzed changes in the population of cotton-top tamarins and its habitat from 2005 to 2012. We used a tailor-made “lure strip transect” method to survey 43 accessible forest parcels that represent 30% of the species’ range. Estimated population size in the surveyed region was approximately 2,050 in 2005 and 1,900 in 2012, with a coefficient of variation of approximately 10%. The estimated population change between surveys was -7% (a decline of approximately 1.3% per year) suggesting a relatively stable population. If densities of inaccessible forest parcels are similar to those of surveyed samples, the estimated population of cotton-top tamarins in the wild in 2012 was 6,946 individuals. We also recorded little change in the amount of suitable habitat for cotton-top tamarins between sample periods: in 2005, 18% of surveyed forest was preferred habitat for cotton-top tamarins, while in 2012, 17% percent was preferred. We attribute the relatively stable population of this Critically Endangered species to increased conservation efforts of Proyecto Tití, conservation NGOs, and the Colombian government. Due to continued threats to cotton-top tamarins and their habitat such as agriculture and urban expansion, ongoing conservation efforts are needed to ensure the long-term survival of cotton-top tamarins in Colombia.
Savage, A., Thomas, L., Leighty, K.A., Soto, L.H., Medina, F.S. (2010) Novel survey method finds dramatic decline of wild cotton-top tamarin population. Nature Communications. 1:30, DOI: 10.1038/ncomms1030
The cotton-top tamarin ( Saguinus oedipus ) is a critically endangered primate, endemic to the tropical forests of Colombia. Population monitoring is essential to evaluate the success of conservation efforts, yet standard survey methods are ineffective because animals fl ee silently before they are seen. We developed a novel technique that combines the use of playbacks of territorial vocalizations with traditional transect surveys. We used remote sensing to identify potential habitat within the species ’ historic range, and visited the 27 % that we could survey safely. Of this, only 99 km 2 was extant forest, containing an estimated 2,045 animals (95 % confi dence interval 1,587 – 2,634). Assuming comparable densities in non-surveyed areas, approximately 7,394 wild cotton-top tamarins remain in Colombia. With 20 – 30,000 animals exported to the United States in the late 1960s, this must represent a precipitous decline. Habitat destruction and capture for the illegal pet trade are ongoing. Urgent conservation measures are required to prevent extinction in the wild.
Miller, L., Savage, A., Giraldo, H. 2004. Quantifying the remaining forested habitat within the historic distribution of the cotton-top tamarin (Saguinus oedipus) in Colombia: Implications for long-term conservation. American Journal of Primatology, 64:451-457.
Landsat Thematic Mapper (TM) data were used to classify forested areas within the historic distribution of the endangered cotton-top tamarin (Saguinus oedipus) in Colombia. This species continues to be threatened by habitat destruction, and we observed a 31% decrease of the forested habitat within the tamarins' historic distribution in 1990–2000. It is estimated that since the establishment of some protected areas for cotton-top tamarins and other native Colombian wildlife (Parque Nacional Natural Paramillo, Santuarío de Fauna y Flora Los Colorados, and Reserva Forestal de Montes de Maria), almost 43% of the original forested area has been lost. Given that the human population of Colombia increases annually by 1.6% [Patel, 2002], it is important to target specific areas for tamarin protection while creating mitigation strategies to compensate for economic growth. The results of this study provide valuable information to assist in the long-term development of effective conservation strategies for this endangered primate.
Return to Top
REPRODUCTION
Wheaton, C.J., Feilen, K.L., Soto, L.H., Medina, F., Emeris, G., Guillen, R., Savage, A. 2022. Seasonality of reproduction in wild cotton-top tamarins (Saguinus oedipus) in Colombia. American Journal of Primatology, 84(2): e23359
Seasonal availability of resources can influence the timing and success of reproduction in primates. This study examines the annual pattern of conceptions, mid-term (13 weeks) and full-term (26 weeks) pregnancy losses, births, and infant survivorship of the Critically Endangered cotton-top tamarin (Saguinus oedipus) at Proyecto Tití's long-term field site in Santa Catalina, Colombia. Using 18 years of behavior, physical exam, and fecal steroid data (N = 51 females, 168 conceptions, 121 liters), fecal estrone conjugate (E1C) and pregnanediol-3-glucuronide (PdG) or progesterone (P4) profiles were examined to investigate the relationships between female reproductive patterns and reproductive success with feeding and climatic data. Cotton-top tamarins demonstrated seasonal patterns of reproduction and pregnancy losses. We found positive correlations between peak conception and nectarivory, births and rainfall, and mid-lactation and frugivory. High body mass mothers reproducing during the peak season were more likely to have infants survive. Overall, these results suggest that tamarins use a “relaxed income-breeder” strategy. Cotton-top tamarins appear to maximize reproductive output at the time of ovulation, and can reduce their investment in the current reproductive effort at many stages. Changes in food availability due to habitat loss and climate change will require the Critically Endangered cotton-top tamarins to alter their reproductive strategies to survive.
Wheaton CJ, Savage A, Soto L, Medina F, Emeris G, Guillen R. 2012. Tolerance may lead to loss of tenure in tamarin societies: Use of non-invasive faecal reproductive steroid monitoring and behaviour observations to reveal reproductive and life-history traits in wild female cotton-top tamarins (Saguinus oedipus) in Colombia. Wien Tierärztl Monat 99 (Suppl 1): 16.
Cotton-top tamarins (Saguinus oedipus) are a critically endangered primate found only in Colombia. Efforts to conserve this species are centered on developing effective management plans that integrate biological and reproductive information regarding population dynamics and factors that influence their survival through pressures of habitat loss. Here, we report 13 years of data on reproductive and life-history traits for wild female cotton-top tamarins (Saguinus oedipus) in Colombia. As part of our long-term field observations on multiple groups, all animals have been individually identified and receive annual exams that include physical and reproductive assessments. Field observations were conducted three to five times weekly by a team of observers to record group composition, inter-group interactions, aggression, births and to collect faecal samples. A total of 8,684 faecal samples were collected from 41 females in 19 family groups over 13 years. Faecal pregnanediol-3-glucuronide and oestrone conjugate (PdG antisera P70; E1C antisera R522-2, C. Munro, UC Davis, CA) analyses were used to assess reproductive condition of the dominant breeding female, daughters and unrelated or immigrating females through changes in reproductive tenure in each group. Average reproductive tenure was 2.4 ± 2.2 years (range 0.2 to 8.2; n=19). We observed 17 occurrences where the dominant female was evicted from the group. In nine of these cases (52.9%), the dominant female was pregnant when evicted. Daughters remained suppressed until 11.1± 0.1 months of age (n=5). However, we observed four instances where mother and daughter were pregnant in the same group. Loss of reproductive tenure was primarily associated with immigration and aggression from unrelated adult females (15 of 17 cases; 88.2 %). Of these, eight of the immigrating females were pregnant. Pressures from habitat loss may increase population density and contact with adjacent groups which may impact lifetime reproductive success in wild female cotton-top tamarins.
Savage, A., Soto, L., Medina, F., Emeris, G., & Soltis, J. (2009). Litter Size and Infant Survivorship in Wild Groups of Cotton-top Tamarins (Saguinus oedipus) in Colombia. American Journal of Primatology, 71, 1-5.
Developing successful conservation programs for endangered primates requires a complete understanding of the factors involved in successful reproduction and causes of infant mortality. In contrast to other nonhuman primates, callitrichids are unusual as litter size can vary between one to five offspring in captivity. Twins tend to be the norm for most callitrichids, but the incidence of triplet and quadruplet births can account for nearly 10–50% of the litters born in captive colonies. Litter size significantly influences the survivorship of infants with twins having a higher survivorship than triplets or quadruplets. In the wild, reports suggest that twin and singleton litters were the norm in most callitrichids studied to date. Incidence of triplet births is very low in the wild, but there have been triplets observed in a Golden Lion Tamarin family where none of the offspring survived and in a Common Marmoset family where evidence was found of survival.
In our study, there were fifty-eight litters from 21 females recorded at the two field sites. Twins were most common (81%), followed by singletons (16%), and we had the first report of triplet births (3%) in wild cotton-top tamarins. Mortality was not uniformly distributed across time with 33% of cases confined to the first week. Determining the sex of the offspring rarely occurred if infants died or were missing from the group, and carcasses of infants were rarely found. Thus, calculating an accurate sex ratio of infants was not possible. However, surviving offspring were comprised of 42 males and 32 females. Determining the immediate cause of mortality was also challenging. Two of the six carcasses that were recovered were too decomposed to determine the cause of death. The remaining four carcasses were from the two observed triplet births and malnutrition was deemed the cause of death.
Fourteen females produced more than one litter. For these females, the mean survival rate to 6 months for first litters was 57%710.3 SEM but the mean across subsequent litters rose to 86%77.6. However, the effects of parity were not as clear. One primiparous female (Nadia) left her family group and assumed the position of the reproductively active female in a neighboring group and she gave birth to twins that survived to 6 months. However, two primiparous females (Milena and Tamara) were identified in the Santa Catalina sample but do not represent ‘‘typical’’ primiparous females as they were in groups containing two pregnant females. Milena successfully cared for her twins until one was observed missing from the group at week 8 and Tamara’s twins were ‘‘kidnapped’’ by her mother and subsequently died. Litter size appeared to influence survival with triplet litters showing a lower infant survival to 6 months (33%) than twins (80%) or singletons (77%), although the number of triplet litters (n52) is too small for statistical analysis.
This study confirms that infant survival to 6 months of age in the wild (78% twins and 88% singletons), gathered from two distinct field sites, and was greater than has been reported in captive colonies. Similar to captive colonies mortality was highest during the first week of life; however, given the low rate of carcass retrieval we cannot determine the cause of mortality and infant mortality associated after 20 days may be related to the development of infant independence. Given that the parity of the majority of our study females could not be determined, we did find that infant mortality was higher in first litters than subsequent litters born to wild cotton-top tamarin females. We were unable to find a significant relationship between infant mortality and group size or the number of adult males in a social group. However, there was a trend toward higher mortality with a shorter inter-birth interval. The number of females that produce multiple litters in 1 year is rare, but infants born out of the normal birthing season may be at risk as food may not be as plentiful for a nursing female.
Return to Top
VOCALIZATIONS
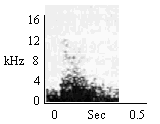
Cotton-top tamarins have an extensive vocal repertoire which is derived from the variation of two basic elements and the sequential combination of those elements (Cleveland and Snowdon 1982). The vocalizations described here were recorded over 18 months from a captive colony in Madison, Wisconsin with a Uher 4200 stereo tape recorder using a Sennheiser MD 441 microphone and either Scotch 209 or 177 low noise recording tape. Recordings were done at 9.5 cm/s. Sound spectrograms were made on a Kay Sonagraph Model 6061 B using a narrow band pass filter (90 Hz) over the frequency range of 160 to 16,000 Hz.
By assessing the structural and behavioral correlates of the vocalizations, 38 distinct sounds or combinations of sounds were identifies. The simplify the description of the repertoire, the sounds were grouped into six major classes based on structure.
Click on the Spectrogram to Hear the Vocalization
Class 1: Single Frequency Modulated Syllables
A. Short Duration Calls – Chirps were categorized according to four structural parameters: presence or absence of stem upsweep, duration, peak frequency, and frequency change. See Table 1 for call parameters.
| Type A Chirp (Mobbing) |
During mobbing behavior, to sudden animated stimuli. By some groups to preferred foods. Rarely given to acoustical stimuli. |
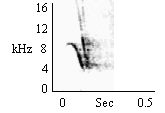
|
| Type B Chirp (Directed Investigatory) |
To human observers or familiar object. |
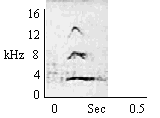
|
| Type C Chirp (Pre-food) |
During approach to food or when individual approaches object that will be hand-held and orally explored. |
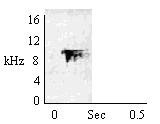 |
| Type D Chirp (Post-food) |
When animal actually possesses food or object. |
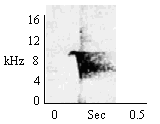 |
| Type E Chirp (General Alarm) |
To sudden visual and acoustic stimuli. To sudden leaping movement by group members if animal startled. |
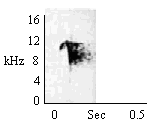
|
| Type F Chirp (Intergroup) |
During intergroup antiphonal calling of Normal Long Calls. To audible outgroup vocalizations. |
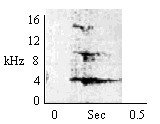
|
| Type G Chirp (Nonspecific Investigatory) |
During relaxed environmental investigation. |
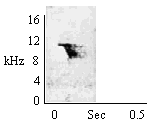
|
| Type H Chirp (Mild Alarm) |
To novel visual stimuli at close proximity. |
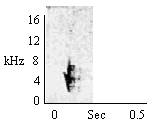
|
Table 1. Parameters differentiating types of chirps.
| Call Type |
Stem Upsweep |
Duration (ms) |
Peak Frequency |
Frequency Change |
| Type A |
Yes |
79.9 ± 29.7 |
8.7 ± 1.7 |
5.9 ± 1.6 |
| Type B |
Yes |
100.0 ± 26.0 |
6.0 ± 1.0 |
2.0 ± 0.6 |
| Type C |
Yes |
25.2 ± 5.4 |
10.4 ± 1.2 |
3.4 ± 1.0 |
| Type D |
Yes |
42.6 ± 10.2 |
8.2 ± 0.7 |
3.3 ± 1.5 |
| Type E |
Yes |
36.9 ± 10.4 |
8.0 ± 1.6 |
3.1 ± 1.2 |
| Type F |
No |
56.0 ± 19.7 |
4.1 ± 0.7 |
1.0 ± 0.4 |
| Type G |
No |
57.9 ± 10.7 |
9.0 ± 1.1 |
6.1 ± 0.7 |
| Type H |
No |
20.8 ± 4.1 |
5.5 ± 0.8 |
2.9 ± 0.8 |
B. Long Duration Calls
| Squeal |
During active physical contact by the passive participant: during wrestle play or face pressing between two individuals. |

|
| Slicing Scream |
Same as Type A Chirp |
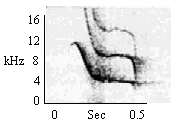 |
Class 2: Pulsed Vocalizations
| Twitter |
Same as Squeal. Results from intensification of Squeal Context. |

|
| Hooked Chatter |
Same as Type D Chirp (food related). As infants approach. |
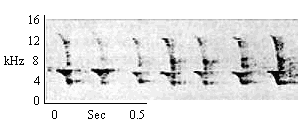 |
| Chevron Chatter |
Towards humans when animal is netted or placed in small cage. Also by some animals in response to human approaching them closely in home cage. |
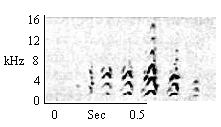 |
| Type F Chirp Trill |
Same as for Type F Chirp. Tilling indicative of a higer state of arousal than Type F Chirp alone. |
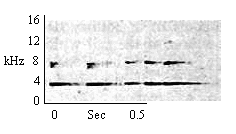 |
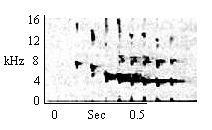
| Type H Chirp Trill |
Same as for Type H Chirp. |
 |
| Type A Trill |
During Antiphonal Call play interactions prior to attempted play mounts. |
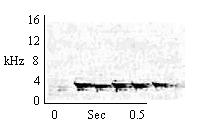 |
| Type B Trill |
By individuals as they approach young infants to retrieve them. Also given by animals carrying young infants when infants move, vocalize or attempt to climb off. |
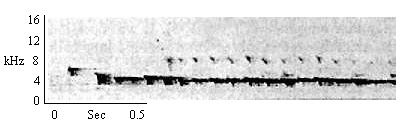
|
| Type C Trill |
Signals the end of a nursing bout, interspersed with Terminally Modulated Whistles. |

|
Class 3: Single Whistles
| Squeak |
In vigilance, foraging and investigatory activities. Increase in rate of emission with increase in arousal. |
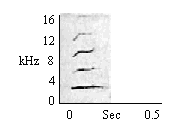
|
| Large Initially Modulated Whistle |
Given greater than 0.6 m from other animals in low arousal situation. Not given during resting, grooming, nursing, except occasionally when animals break contact. Occurs during Antiphonal Call play |
 |
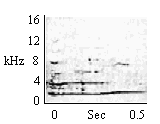
| Small Initially Modulated Whistle |
Same as Large Initially Modulated Whistle except given at distances less than 0.6 m from other group members. |
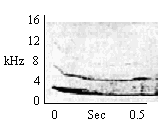 |
| Terminally Modulated Whistle |
By individuals in resting contact, during grooming, by mothers during nursing, by parents and siblings retrieving and carrying infants. |
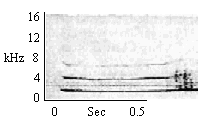 |
| Flat Whistle |
Given following Type F, H or E Chirp Trills |
 |
Class 4: Multiple Whistle Calls
| Ascending Multi-Whistle (Intragroup) |
By animals in huddles, by mothers nursing infants. |
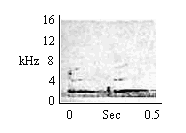
|
| Descending Multi-Whistle (Intragroup) |
Same as Ascending Multi-Whistle and Terminally Modulated Whistle. More commonly given to infants. |
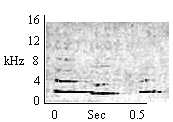 |
| Partial Quiet Long Call (Intragroup) |
By mothers nursing infants. By subadults during close huddle contact and pauses during wrestle play. |
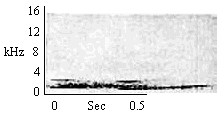 |
| Rapid Whistles (Intragroup) |
Before an approach to mount during Anti-phonal Call play. |
 |
| Quiet Long Call(Intergroup) |
To audible Single Whistles, Chevron Chatters, Type H Trills, Twitters, Long Calls from unfamiliar animals. Occasionally given to no obvious stimulus during group huddles. |
 |
| Normal Long Call (Intergroup) |
To distant non-group Combination Long Calls, Normal Long Calls, Type F Chirps. During isolation. |
 |
Class 5. Combination Vocalizations
| Small Modulated Multi-Level |
Same as Small Initially Modulated Whistle |
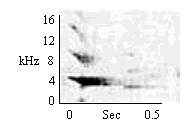
|
| Large Modulated Multi-Level |
Same as Large Initially Modulated Whistle |
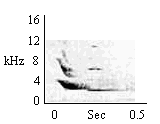 |
| Combination Long Call |
During play, isolation or when socially disturbed. When animal not in contact. |
 |
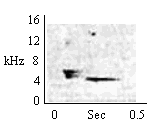
| Inverted U + Whistle |
In alarm to faint acoustical stimuli after an alert has been signaled. |
 |
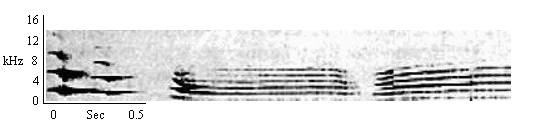
| Type F Chirp + Whistle |
By individuals less confident than when giving Normal Long Calls. As response to Combination or Normal Long Calls or non-group Type F Chirps. In isolation. |
 |
Class 6. Noisy Calls
| Squawk |
By recipient of aggressive behavior. In some animals as an appeasement gesture, by others as an invitation for grooming. |

|
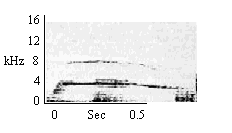
| Scream |
When animals attempt to steal food. |
 |
| Sneeze |
After eating, drinking water, sniffing or as result of rubbing nose on substrate or with hand. |
|
Return to Top